Thismia hillii (Cheeseman) N. Pfeiffer
Cheeseman (1908) described Bagnisia hillii from specimens collected by Mr. H. Hill on the northern Island of New Zealand, illustrated in 1914 by drawings of M. Smith in the "Illustration of the New Zealand Flora" (Vol. II, Plate 191 A, see under 'Media'). He follows Engler (1889) on the synonymy of Geomitra with Bagnisia (both coined by Beccari 1877) the synonymy of which has already been strongly advised ("Genus forte melius Bagnisiae adjungendum") by Bentham and Hooker (1883). Hence, Mr. Hills finding became included in Bagnisia, and Cheeseman mentions Geomitra only in parantheses. Pfeiffer (1914), in an enumeration of existing Thismias, changed Bagnisia hillii in Thismia hilli with the preceding annotation " The Geomitra and Bagnisia divisions had been described as separate genera by earlier workers, but have recently been included in the genus Thismia." Unfortunately, she did not mention her reference, but it is likely that she meant the works of Ernst and Bernard (1910-1912) and J. J. Smith who indeed can be quoted: "I believe that Baron v. Müller was right in uniting the genus Bagnisia Becc. (with Geomitra Becc. and Sarcosiphon Bl.) with Thismia Griff." (Smith 1909, translated from German). In contrast, the rather splitting taxonomist Schlechter (1921) preferred the older name Sarcosiphon from Blume (1849) for those Thismieae having the inner perianth lobes bent upwards and connected above the flower tube (building a 'mitre'), and transferred Bagnisia hillii to Sarcosiphon hillii. Cheeseman (1908) obviously did not compare his new species with Thismia rodwayi (von Mueller 1890), the author of which preferred the wider circumscription of Thismia, just like it is the notion today. For Cheeseman the genus Thismia was characterized by six free petal lobes, not matching with Mr. Hills specimen. However, Thismia rodwayi and Bagnisia hilli are so similar, that most following scientists agreed on their identity (e.g. Jonker 1938, Coleman 1941, McLennan 1958, Campbell 1968, Maas et al. 1986, Roberts et al. 2003), accepting an extension of its geographical distribution to New Zealand. In fact, anatomical and mycorrhizal investigations done by Ella Campbell (1968) were based on Thismia hilli rather than T. rodwayi.
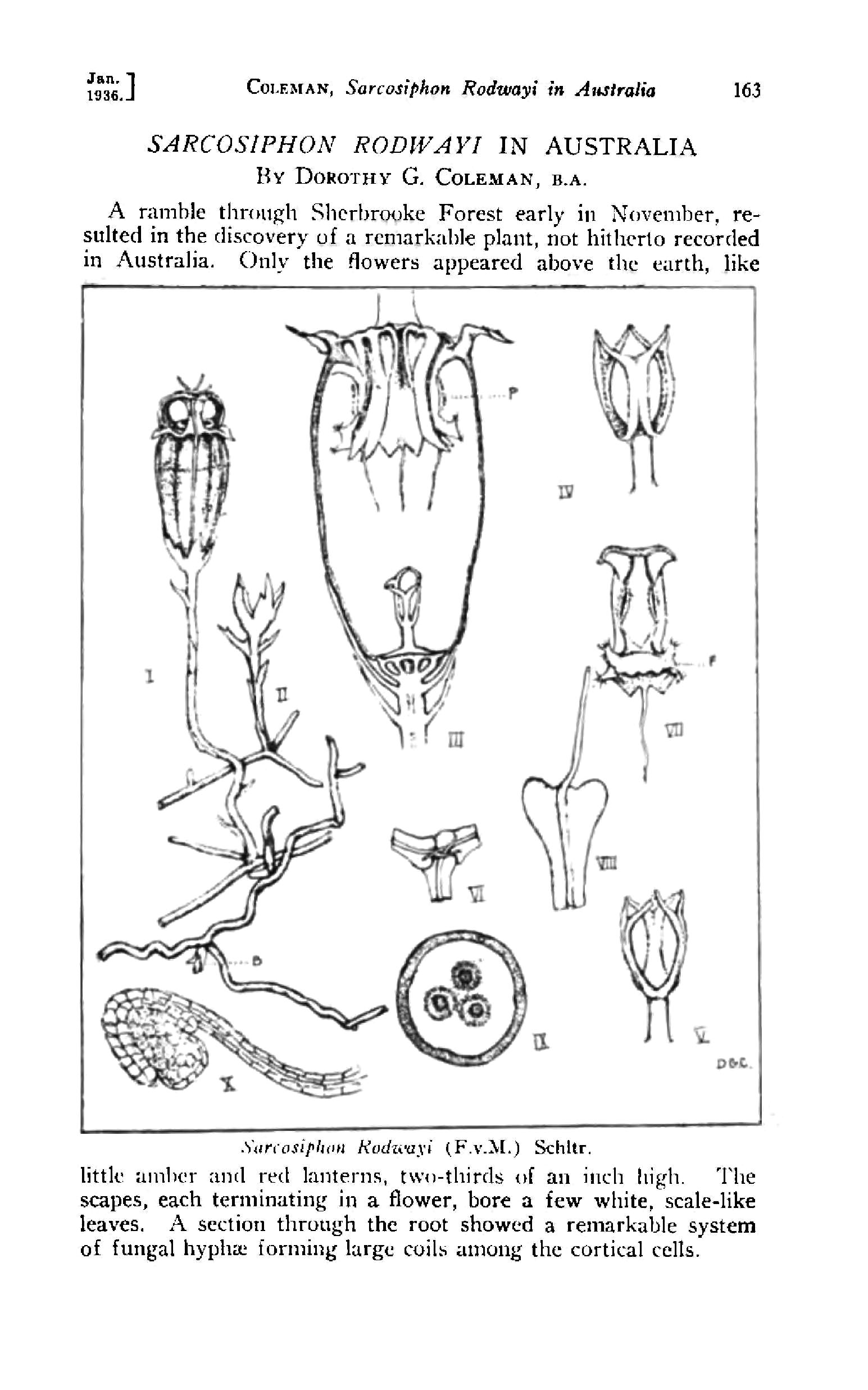
It was not until 2014, when Merckx and Smets (2014) made molecular analysis on many Thismiaceae including those from the section Rodwaya, namely Thismia clavarioides, T. huangii and T. rodwayi from three locations (Tasmania, Victoria/Australia and New Zealand). Surprisingly the specimen from Tasmania and Victoria of T. rodwayi were closer to T. clavarioides than to their 'conspecific' specimen from New Zealand. The authors had two options: either unifying T. clavarioides with all T. rodwayi, or resurrecting Thismia hillii (= T. rodwayi specimen from New Zealand) in order to keep the monophyly of T. rodwayi. Since morphologically T. clavarioides is clearly different from T. rodwayi the former option would have been hard to justify. Hence, Thismia hillii is now considered as an own species again. Retrospectively, the drawings provided by the describing authors as well as the one from Coleman (1936) indeed show differences! The stigma in T. hillii is tripartite and its lobes are rather rounded, whereas in T. rodwayi they seem to be split in six pointed parts, perhaps in pairs united at their tips (in the drawings of Coleman 1936). Moreover, the outer perianth lobes in T. rodwayi are rather triangular, those of T. hillii more elongated, and the flower subtending bracts seem to be serrate in T. rodwayi but entire in T. hillii (see plates under 'Media'). It would be interesting to know if these differences indeed can be found in the herbarium specimen collected either in New Zealand or Australia/Tasmania.
")
 after Coleman (1936)")
 after Cheeseman (1908) in Illustrations of the New Zealand Flora, Vol. 2, t191")
{kind=link}
{kind=link}
{kind=link}